
Genetik, Paylaşılmayan Çevre ve Paylaşılan Çevre
Paylaşılmayan Çevre nedensel midir?

Inquisitive Bird (IB), geçtiğimiz yıl genetik ve paylaşılan çevre etkilerini inceleyen bir yazı yazmıştı. Paylaşılan Çevre üzerinden ailelerin çeşitli özelliklerde ve davranışlarda ne derecede fark yarattığını gösteren bu çalışma genetik çalışmalarına bakılınca her şeyin genetik olduğunu ve ailelerin önemsiz olduğu algısını yıkıyor.
Not: Kaynakları hiperlink olarak koyacağım. Altı çizili gördüğünüz çalışmaya tıkladığınızda doğrudan kaynağa gideceksiniz, ayrı bir kaynakça yazmayacağım. Örneğin: Knopik vd. (2018)

Öncelikle, bu kavramları açıklamak için Knopik vd. (2018) tarafından yazılan Davranışsal Genetik kitabını kullanacağım:
Paylaşılan çevre etkileri (kısaca C), aile üyelerini birbirine benzeten genetik dışı tüm etkileri ifade eder. Bu; mahalle, ebeveyn eğitimi ve ebeveynlik tutumları veya ev içindeki çatışma ya da ev içindeki kaos gibi ailevi faktörleri içeren geniş bir yelpazeyi kapsayabilir. Bu faktörlerin "paylaşılan çevre etkisi" sayılabilmesi için, yalnızca aynı evde yaşayan bireyler arasında daha fazla benzerliğe yol açması ve genetik yakınlığa bağlı olarak değişiklik göstermemesi gerekir. Diğer bir deyişle, çift yumurta ikizleri en az tek yumurta ikizleri kadar birbirine benziyorsa ve bu benzerlik göz ardı edilemeyecek düzeydeyse, paylaşılan çevre etkileri önemlidir. Benzer şekilde, aralarında genetik bağ olmayan "evlatlık" kardeşler, "genetik" kardeşler kadar birbirine benziyorsa, paylaşılan çevresel etkilerin varlığına işaret edilir.
Paylaşılmayan çevre etkileri (kısaca E), ölçüm hatası da dahil olmak üzere, aile üyeleri için bağımsız (veya birbiriyle ilişkisiz) olan tüm genetik dışı etkilerdir. Aynı evde yaşayan tek yumurta ikizleri tüm genlerini ve çevrelerini paylaştıkları için, tek yumurta ikizi çiftleri arasındaki farklılıkları açıklayabilecek tek şey paylaşılmayan çevre etkilerdir. Paylaşılmayan çevre etkisi kaynakları arasında; ebeveynlerin farklı muamelesi gibi aile içi deneyim farklılıkları veya farklı arkadaşlara sahip olmak gibi aile dışındaki farklı deneyimler yer alır.
Bu modeller haricinde de bizim kısaca bahsettiğimiz genetik etkiler vardır, kısaca A. Böylece modelin kısaltması ACE’dir. Bu modeli ekonometristler, davranış genetiği uzmanları veya epidemiyoloji uzmanları kullanır.
Paylaşılan Çevreyi Anlamak
Inquisitive Bird (IB), bu kısa yazısında davranışsal genetik (Behavioral Genetics) alanındaki sonuçların yanlış yorumlanmasına dikkat çeker. Bunlardan biri, bir özelliğe ait varyansın "küçük" bir kısmının paylaşılan çevre (C) tarafından açıklanmasının, C'nin önemsiz olduğu ve dolayısıyla göz ardı edilebileceği anlamına gelmesidir. Bu yanılgıyı açıklamanın kolay yolları olduğunu düşünüyorum. Örneğin, tipik bir IQ dağılımının varyansının (toplam örneklemin değil) en alttaki %5, %10 ve %20'si çıkarıldığında dağılım ortalamasına ne olduğuna bakalım.
Danimarka’da 1 milyondan fazla erkeğin verileriyle yapılan kapsamlı bir çalışma, düşük IQ seviyesinin (özellikle 80 ve altı) okul bırakma, işsizlik, sosyal yardıma bağımlılık ve düşük gelir gibi başarısızlık göstergeleriyle çok güçlü bir korelasyon içinde olduğunu kanıtlamaktadır (Hegelund vd. 2018). İstatistiksel olarak varyansın sadece %5-10’unu etkileyen çevresel iyileştirmeler bile, bireyleri bu kritik risk barajlarının üzerine çıkararak onların sosyal ve ekonomik kaderini kökten değiştirebilir; bu da bize "küçük" görünen çevresel etkilerin, aslında toplumsal müdahale ve eğitim planlaması için en stratejik olduğunu göstermektedir.
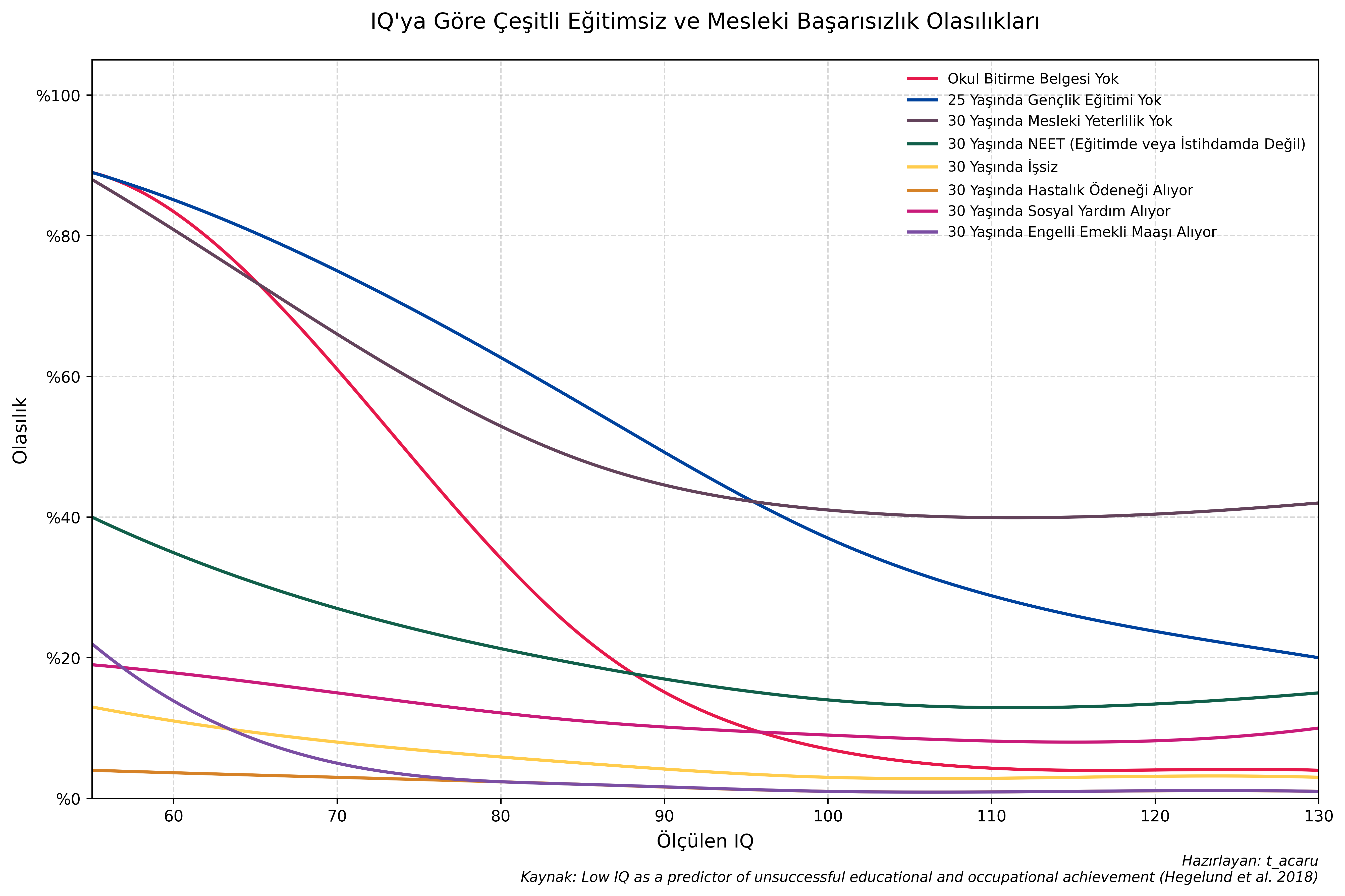
Bunu gerçekleştirmenin sosyal etkileri oldukça kayda değer olacaktır; benzer bir durumu suç verileri için de geçerlidir (Schwartz vd. 2015; Frisell vd. 2012). Küçük etkilerin çok şey ifade edebileceğini göstermenin pek çok başka yolu da vardır: Abelson Paradoksu. Örneğin %40'tan az etkili sıtma aşıları çocuk ölümlerini %13 azaltmıştır. Not: Scott Alexander’ın bu konudaki yazısını okumanızı tavsiye ederim.
İkinci olarak Paylaşılan çevrenin (C) küçük görünen etki büyüklükleri, sırf "küçük görüldüğü" için bir yok sayılamaz. Etkiler genellikle standartlaştırılmış terimlerle karşılaştırılır, oysa doğru bir karşılaştırma için farklı varyans bileşenlerinin etkilerini "standartlaştırılmamış" terimlerle anlamak gerekir. Örneğin, klasik ikiz çalışmalarındaki eklemeli genetik etkiler (A) bir özelliğin varyansının %80'ini açıklıyorsa, bunların o özellik için çevreye kıyasla dört kat değil, neden sadece iki kat daha önemli olduğunu açıklar. Eğer genler çevrelerden gerçekten dört kat daha önemli olsaydı, genler varyansın %80'ini açıklarken çevreler sadece %5'ini açıklardı. Bu durum, etkileri anlamak için çok daha sezgisel bir çerçeve sunar; sonuçta 0,80 ile 0,20 arasındaki fark, 0,8944 ile 0,4472 arasındaki farktan çok daha büyüktür.
Bunu daha geniş bir şekilde açıklayayım. Davranışsal genetik modellerinde genetik (A) veya paylaşılan çevre (C) gibi bileşenler genellikle varyansın yüzdesi (R kare) olarak ifade edilir. Varyans, birimlerin karesi alınarak hesaplanan bir istatistiktir. Bu durum, büyük etkileri çok daha büyük, küçük etkileri ise çok daha önemsizmiş gibi gösterir. Örnek: Bir özelliğin varyansının %80'i genetik (A=0,80), %20'si çevresel (E=0,20) olsun. Varyans Oranı: 0,80/0,20=4. Bu rakama bakınca "Genler çevreden 4 kat daha önemli" dersiniz; lâkin bir etkenin o özellik üzerindeki gerçek "doğrusal gücünü" veya "önemini" anlamak için varyansın karekökünü alarak etki katsayısına (r) bakmamız gerekir. Bu, değişkenleri "kareler" evreninden çıkarıp, normal birimler evrenine geri döndürür.
0,80’nin karekökü: 0,894; ,20’nin karekökü 0,447. Gördüğünüz gibi, varyans tablosunda aralarında "uçurum" (4 kat) var gibi görünen genetik ve çevre, aslında etki gücü bakımından birbirine çok daha yakındır (sadece 2 kat fark).
IB'nin makalesinin geri kalanı, çeşitli sosyal özellikler üzerindeki paylaşılan çevre (C) etkisine dair en iyi kanıtların bir incelemesiydi; eğitim, gelir, suç ve servet için genetik etkinin (A), paylaşılan çevreden (C) sırasıyla 1,4-2,1, 1,7-2,3, 2 ve 1,3 kat daha fazla olduğu sonucuna varıyordu. Ek olarak, genetik faktörler genel olarak paylaşılan çevreden daha önemli olsa da kardeşleri birbirine ne daha benzer ne de daha az benzer kılan faktörler olan paylaşılmayan çevre (E), genellikle en önemli faktördü. Bu durum eğitim veya suç için geçerli değildi; lâkin gelir ve servet için geçerliydi. Dolayısıyla, sistematik faktörlerin (A ve C) ötesinde, rastgelelik ve kardeşlerin farklı tepki verdiği çevreler de önem taşımaktadır.
Standart ikiz modelinde paylaşılan çevre (C), teoride, kardeşleri birbirine daha benzer kılan ve genlerin eklemeli etkileri dışındaki tüm faktörlerdir. Fakat, C olarak adlandırdığımız şey, aslında baskın genetik etkiler (D) veya eş seçimindeki benzerliğin (assortative mating) bir sonucu olduğunda bu teori çöker. Dahası, bir faktörün kardeşleri birbirine daha benzer kılıyor olması, o faktörün mutlaka tüm aile üyelerini birbirine benzettiği anlamına gelmez. C, her zaman ebeveynlerden dikey olarak aktarılan bir sürecin ya da aileye has özel bir durumun sonucu değildir; bazen sadece kardeşlerin paylaştığı ortamdan kaynaklanır. Hatta bazen C, kardeşlerin genel olarak paylaştığı bir şey bile değil, sadece ikizlerin paylaştığı bir durumdur. Wolfram ve Morris (2023) çalışmalarında eğitim düzeyi için bunun bir örneğini sunmuşlardır. Standart ACE modellerinde eğitim başarısını %43 oranında belirlediği sanılan paylaşılan çevre bileşeni, daha gelişmiş modellerle incelendiğinde üç farklı parçaya bölünmektedir: Ebeveynlerin benzer eğitim seviyelerine sahip kişilerle evlenmesinden kaynaklanan eş seçimi etkisi, sadece aynı yaştaki ikizlerin paylaştığı teknik benzerlikler ve gerçek aile ortamı. Bu ayrıştırma yapıldığında, tüm kardeşleri birbirine benzeten asıl aile etkisinin aslında %43 değil, sadece %10 gibi çok daha mütevazı bir seviyede olduğu; buna karşın genetik faktörlerin payının %34'ten %51'e çıktığı görülmektedir. Sonuç olarak bu tablo, "aile etkisi" olarak etiketlediğimiz pek çok unsurun aslında ya genetik bir yansıma ya da sadece ikiz olmaya özgü geçici durumlardan ibaret olduğu tartışmasına neden olmaktadır.
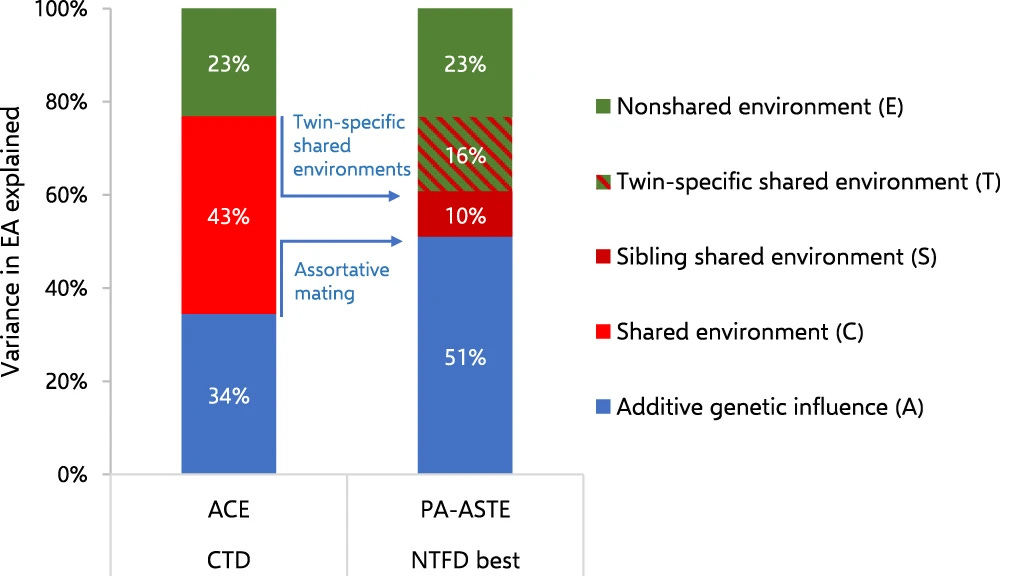
C (paylaşılan çevre), farklı yaşlarda aynı anlama (aynı şey, olgu, fenomen) bile gelmeyebilir. Örneğin, boy özelliği için çocuklukta görülen C muhtemelen gerçekten aile ortamıyla ilgiliyken; yetişkinlikte muhtemelen daha çok eş seçimindeki benzerlikle (assortative mating) ilgilidir. Bu durum ebeveynlerin, ailelerin, kardeşlerin veya ikizlerin üzerinde pek kontrol sahibi olabileceği bir şey değildir.
Varyans Ayrıştırması ve Nedensellik
İkiz ve aile tasarımlarında tanımlanan varyans bileşenleri, akrabalık bağlarındaki varyasyonlar aracılığıyla tanımlandıkları için aslında nedensel niceliklerdir. Bu modellerle nedensel bir tanımlama elde etmek için gereken tek şey, çevrenin, genetik olarak daha benzer olan bireyler için özellikleri anlamlı derecede etkileyecek şekilde sistematik olarak daha benzer olmamasıdır. Ölçülen bu nedensel güce rağmen, yüksek kalıtım oranlarını 'çevresel müdahale imkansızdır' şeklinde yorumlamamak gerekir. Zira evlat edinme çalışmalarından elde edilen nedensel tahminler ile varyans analizleri arasındaki ilişkiyi anlamak, iki temel zorluk nedeniyle karmaşıktır. Bunlardan ilki, görünüşte devasa olan çevresel etkilerin bile aslında çok yüksek kalıtım oranlarıyla bir arada var olabilmesidir. Tıpkı Jensen'in IQ verilerinde veya son yüzyıldaki boy artışı örneğinde gösterdiği gibi; bir özelliğin bireyler arasındaki farklarının %80'i genetik olsa bile, o özelliğin genel ortalaması çevresel müdahalelerle dramatik bir şekilde yukarı taşınabilir. Jensen, Skodak ve Skeels'in (1949) ünlü evlat edinme çalışmasını inceleyerek bunu gösterir. Basitçe, biyolojik annelerinin IQ ortalaması 85.7 olan çocuklar, üst sosyo-ekonomik düzeydeki aileler tarafından evlat edinildikten sonra 106 IQ ortalamasına ulaşmışlardır. İlk bakışta bu 20 puanlık devasa artış, zekânın yüksek oranda kalıtımsal olduğu bilgisiyle çelişiyor gibi görünebilir. Jensen, basit bir poligenik model kullanarak bu artışın %70-80 gibi yüksek bir kalıtım oranıyla tam bir uyum içinde olduğunu göstermiştir. Matematiksel olarak:
M (Popülasyon Ortalaması): 100.
P (Ebeveyn Ortalaması): Skodak ve Skeels örneğinde biyolojik annelerin IQ ortalaması 85.7'dir. Babalar bilinmediği için popülasyon ortalaması (100) kabul edilirse, ebeveyn ortalaması P=(85.7+100)/2=92.85 olur.
hn2 (Kalıtım Derecesi): Jensen burada 0.706 değerini kullanır.
O=100+0.706(92.85−100)≈95. Yani, bu çocuklar ortalama bir çevrede büyüseydi, genetik mirasları gereği 95 IQ almaları bekleniyordu. Fakat çocuklar evlat edinildikleri üst düzey evlerde 106 IQ ortalamasına ulaştılar. Aradaki 11 puanlık fark, çevrenin nedensel etkisidir. Peki, %80 kalıtım varken 11-20 puanlık bir artış nasıl "küçük" bir çevresel etkiyle açıklanır? Jensen bunu varyansın parçalarına ayırarak gösterir:
Toplam Varyans: IQ testlerinde standart sapma 15, yani varyans 152=225’tir. Kalıtım (h2=0.80): Varyansın %80'i genetikse, çevrenin payı %20'dir (1−0,80=0,20). Çevresel Varyans: 225×0,20=45. Çevresel Standart Sapma (σe): 45 ≈ 6,7.
Çevresel koşullardaki her 1 standart sapmalık iyileşme (örneğin daha iyi eğitim) bireyin IQ'sunu 6.7 puan yukarı taşır. Skodak ve Skeels çalışmasındaki çocukların ulaştığı 106 IQ, genetik beklentinin (95) yaklaşık 11 puan üzerindedir. Jensen, evlat edinen ailelerin sosyo-ekonomik ve kültürel düzeyinin genel popülasyondan yaklaşık 1.5 ila 2 standart sapma yukarıda olduğunu belirtir. Yani matematik tam olarak birbirini tutmaktadır: Çok yüksek kalıtım oranları altında bile, çevreyi 1.5-2 birim iyileştirdiğinizde, toplumun veya bireyin başarısını 15-20 puan artırabilirsiniz.
İkincisi, farklı araştırmalarda farklı modeller üzerinden genellikle farklı şeyler ölçüldüğü için, bu farklı tasarımlar yapay nedenlerle sıklıkla farklı sonuçlara varmaktadır. Davranışsal genetik modellerini kullananlar genellikle örtük/gizil değişken (latent variable) kavramlarını ve bunların önemini kavrarlar; bu nedenle, diğer her şey eşit olduğunda, sadece IQ puanlarını modellemek yerine genel zekâ (g) gibi örtük bir değişkeni modelleme ihtimalleri daha yüksektir. Bu çok önemli olabilir çünkü IQ puanlarındaki evlat edinme kazanımları g-dışı faktörlerden kaynaklanıyor olabilir; bu da potansiyel olarak bu kazanımların önemini sınırlar ve kazanımları, gruplar arasında kesitsel olarak var olan farklardan farklı bir şekilde yorumlanabilir hale getirir.
Buna benzer bir sorun, bazı işlem/treatment etkilerinin psikometrik yanlılığın bir sonucu olmasıdır; test puanları üzerindeki işlem/treatment etkilerinin, test performansının temelini oluşturan yetenekleri geliştirmek yerine yalnızca insanları test çözmede daha iyi hale getirmesi durumunda olduğu gibi. Muamele/işlem etkisi bir grubun puanlarını yükseltip diğerininkini yükseltmediği ve grupların başlangıçta tarafsız bir şekilde karşılaştırılabildiği durumlarda bunu anlamak kritiktir çünkü bu durum, müdahale sonrası karşılaştırmanın yanlı (biased) hale geldiği anlamına gelir.
Ekonometrik veya epidemiyolojik tahminlerin davranışsal genetik modellerden elde edilenlerle doğrudan karşı karşıya getirildiği çok fazla ampirik örnek yoktur; fakat var olan az sayıdaki örnek birbirleriyle yakından örtüşme eğilimindedir. Dikkat çekici bir örnek, Sacerdote'un (2007) Koreli evlatlıklar üzerine yaptığı çalışmadır. Bu çalışma gerçekten iyi bir evlatık edinme çalışmasıdır çünkü ebeveynlerin hangi Koreli çocuğu alacakları konusunda hiçbir seçim şansı yoktur; bu da yerleştirme/seçilim konusundaki endişeleri ortadan kaldırır ve Koreli evlatlıklar arasında yerleştirme yaşı ile zekâ arasında neden bir ilişki olmadığını potansiyel olarak açıklar.
Sacerdote’un çalışmasında, yüksek eğitimli ve küçük bir aileye yerleştirilmenin tahmini etkisinin; daha yüksek eğitim alma (örneğin lise bitirince 12 yıl, lisans bitirince 16 yıl, YL bitirince 18 yıl gibi), herhangi bir üniversiteden mezun olma olasılığının artması ve seçkin bir üniversiteden mezun olma olasılığının yükselmesi şeklinde eğitim başarısında önemli bir artışa yol açtığı bulunmuştur (“Evlat Edinme Muamele/İşlem Etkisi”). Evlat edinen ailedeki paylaşılmayan çevre (C) kalitesinde bir standart sapma yukarı çıkmanın etkisi de buna oldukça benzer çıkmıştır (“Varyans Ayrıştırma Etkisi”).
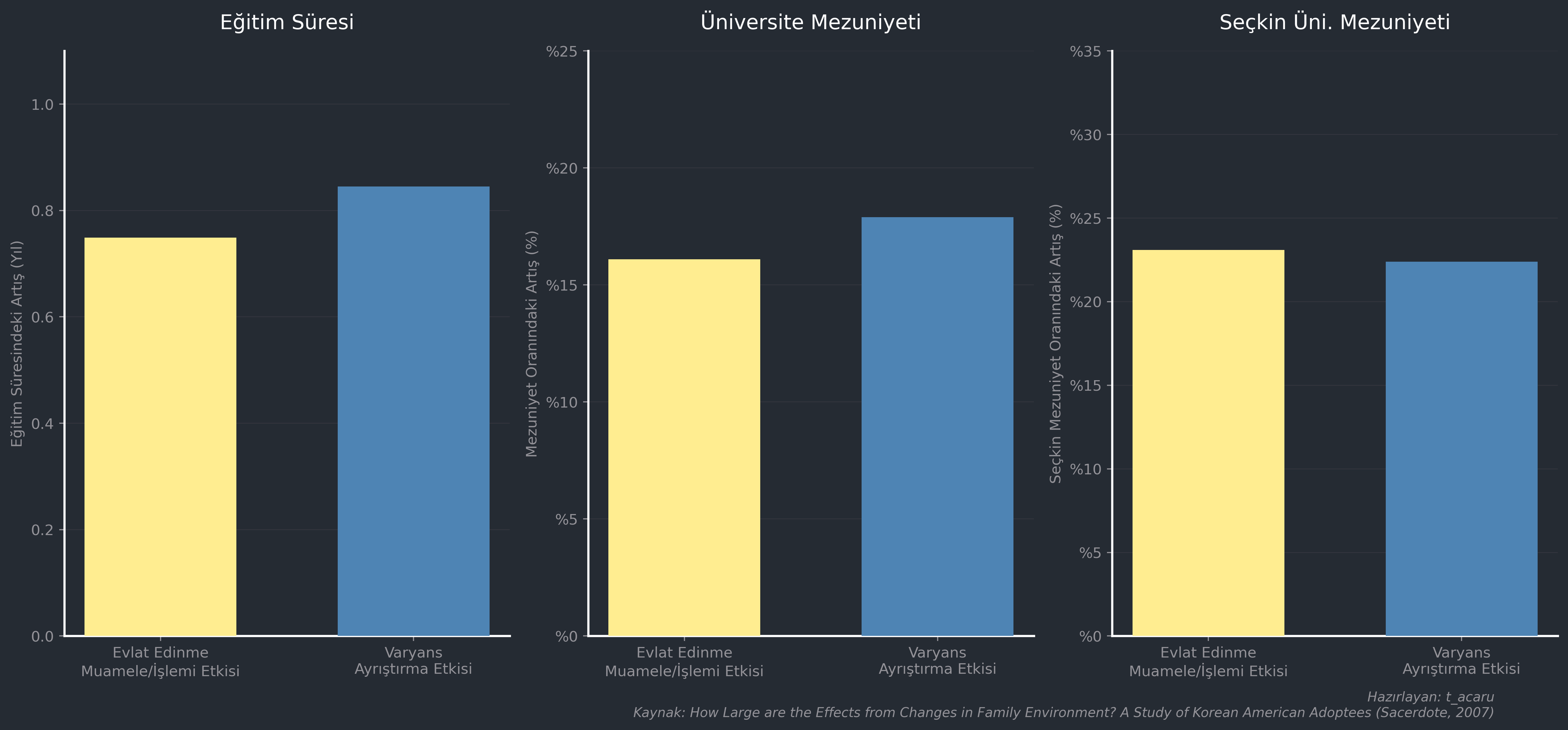
İkinci etki (varyans ayrıştırma etkisi), evlat edinme işlem/muamale etkisinden tutarlı bir şekilde daha büyüktür fakat büyüklük olarak ona benzerdir. Bunun muhtemel nedeni, evlat edinme etkisinin çevre kalitesini az çok kabul edilebilir bir düzeyde temsil etmesi; varyans ayrıştırma etkisinin ise sadece ebeveynin sosyoekonomik durumuna odaklanmak yerine, kardeşleri birbirine benzeten çevre özelliklerini örtük (latent) düzeyde ve kapsamlı bir şekilde endekslediği için hedefi tam on ikiden vurmuş olmasıdır. Davranışsal genetik tahminleri, muhtemelen çevrenin genişliğini daha dikkatli bir şekilde ölçtükleri için daha büyük bir çevre etkisiyle tutarlılık göstermiştir.
Bu durum birkaç noktaya parmak basıyor. Her şeyden önce, bu modellerin ne kadar iyi çalıştığını gösteriyor. İkinci olarak, bu evlat edinme senaryosundaki C (paylaşılan çevre) bileşeninin özelliklerine dair bize birkaç şey söylüyor: Bu bileşenin, kardeşlere özgü bir şeyden ziyade, doğrudan aile ortamının kalitesini endekslediği açıkça görülüyor.
Buradan yola çıkarak davranışsal genetik modellerine dair bir bakış açısı da şudur: Bu modeller, "işlem/muamele etkilerini" (treatment effects) hızlı bir şekilde elde etmek için harika yollar sunar. Örneklemdeki akrabalık derecelerinin çeşitliliği sayesinde, her türlü durum için rastgele tanımlanmış varyans bileşenlerinin nedensel tanımlaması mümkün olduğundan, uygulamacılar bu modelleri oldukça ileri taşıyabilirler. Örneğin, verilerinizde kuzenler varsa, örnekleminizde evlatlıklar olmasa bile "C" bileşeninin evlat edinme çalışmalarındakiyle aynı şekilde tanımlanmasını kolayca sağlayabilirsiniz.
Gen-Çevre Etkileşimi ve Modeller
İnsanlar genellikle, paylaşılan çevrenin (C) neden nispeten küçük bir paya sahip olduğunu açıklamak için gen-çevre etkileşimlerine sığınmak isterler. "A faktörü (genetik) aslında saf bir A faktörü değil, gen-çevre etkileşimli bir AxC faktörüdür" gibi iddialarda bulunurlar. Büyük ölçekli hiçbir çalışmada bu tür bir duruma dair kanıt bulunamamıştır. Bu konuyla ilgili daha çok çalışma gereklidir, özellikle IQ alanında.
Sadece ikizlerle çalışıyorsanız, bir AxC etkileşim faktörünü analitik olarak tanımlayamayabilirsiniz. Ancak bu bir sorun değildir; çünkü yeterli istatistiksel güce sahipseniz, ikiz modelinizden elde edilen faktör skorlarının dağılım özellikleri (momentleri), bu faktörlerdeki varyansın etkileşimli olup olmadığını ortaya çıkaracaktır. Elinizde ikizlerden daha geniş bir veri seti varsa, bu etkileşimli faktörlerin analitik olarak tanımlanması zaten mümkündür. Farklı insan özellikleri üzerinde epistasi (genler arası etkileşim) ve baskınlık etkilerini araştıran Hill, Goddard ve Visscher (2008) da esasen şu sonuca varmışlardır: "Genler düzeyindeki etkileşimlerin, varyans (gözlemlenen farklılıklar) düzeyinde kayda değer bir etkileşim yaratması pek muhtemel değildir." Aynı durum muhtemelen gen-çevre etkileşimleri için de geçerlidir: Bazı etkileşimler mevcut olsa bile, genel olarak bunlar özelliklerdeki toplam varyansa çok az katkıda bulunur.
Gen-çevre etkileşimlerini test etmenin bir başka yolu da evlat edinme verilerini kullanmaktır. Halpern-Manners vd (2020) bunu incelemiştir.. Biyolojik ve evlat edinen ebeveynlerin eğitim düzeylerine dair verilere sahiptiler ve bu düzeyleri evlatlıkların üç farklı Woodcock-Johnson (zeka ve yetenek) puanı üzerinden analiz ettiler. Bulgularına göre; evlat edinme etkileri "değer aralığı kısıtlaması" (range restriction) nedeniyle azalmamıştı ve doğum sonrası temasın etkiler üzerinde düzenleyici (moderator) bir rolü yoktu. En önemlisi, evlat edinen ve biyolojik ebeveyn etkilerinin toplamı, evlat edinilmemiş normal örneklemdeki biyolojik ebeveyn etkisine eşitti. Diğer bir deyişle, ebeveynlerin biyolojik ve çevresel etkileri etkileşimli değil, eklemeliydi (additive).
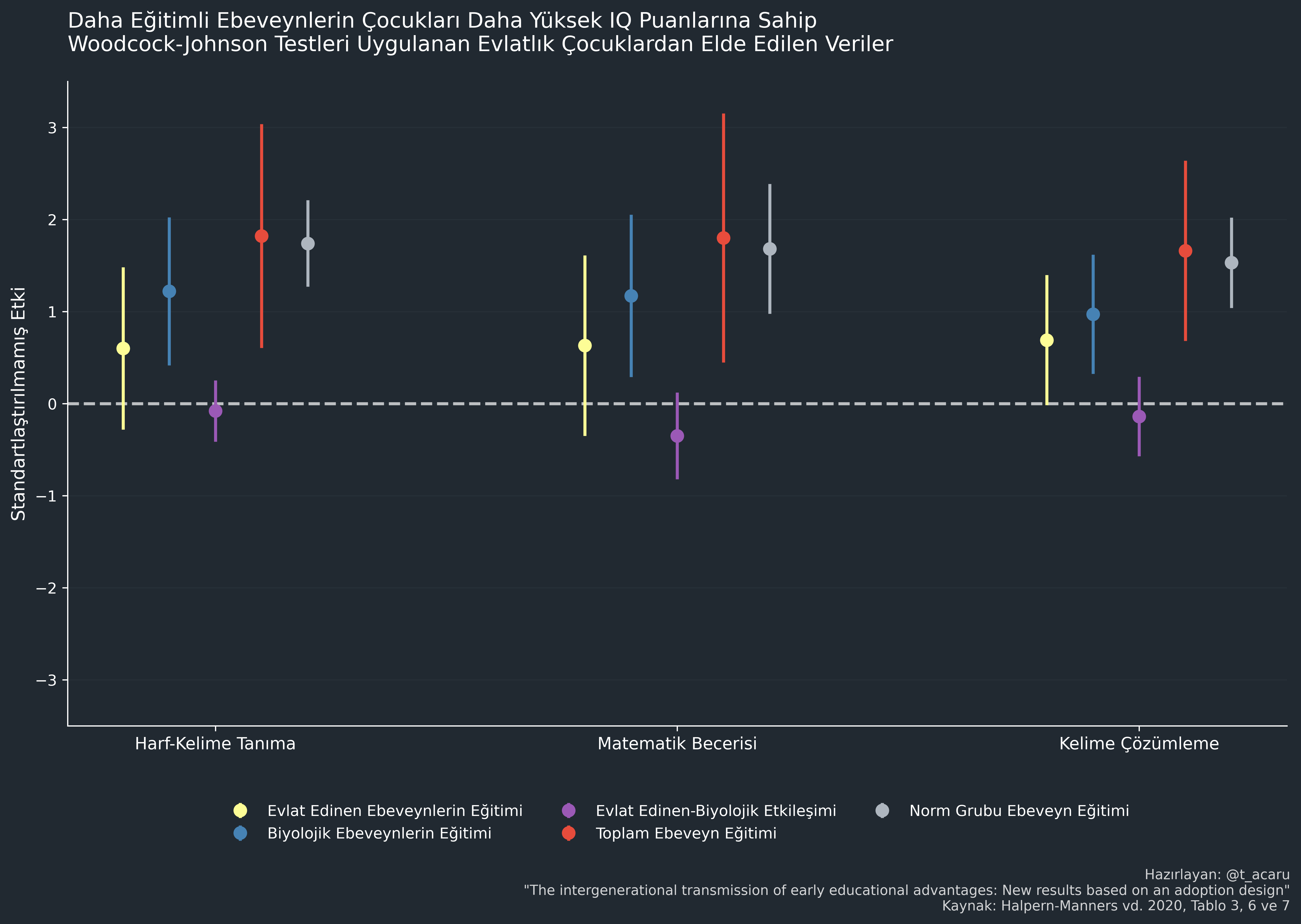
Paylaşılan Çevre Hakkında Öğreneceğimiz Çok Şey Var
Birçok fenotip için paylaşılan çevre (C) tahminlerinin farklı ikiz ve aile modellerinde birbiriyle örtüştüğünü biliyoruz; lâkin bazen örtüşmeyebilirler. Frisell vd (2011) ; klasik ikiz tasarımı, evlatlık-ebeveyn, evlatlık-kardeş ve klasik ikiz benzeri biyolojik kardeş tasarımlarını karşılaştırdıkları çalışmada, şiddet içeren suç işleme eyleminin kalıtım derecesi tahminlerinde, evlatlık verisi kullanılan ve kullanılmayan tasarımlar arasında biraz farklı sonuçlar elde edilmiştir.

İkiz ve biyolojik kardeş modelleri için yapılan ölçümler ayırt edilemez derecede benzerdir,ama C varyansı ("Aile ortamı") için yapılan ölçümler evlatlık (adoptee) modellerinde daha düşük çıkmıştır. Güven aralıkları (CI) oldukça genişti ve negatif değerlere ulaştıklarında yazarların bunları sıfırda sabitlemeyi seçmesi hatalıydı; yine de en yüksek istatistiksel güce sahip model (biyolojik kardeş modeli) her iki evlatlık modeliyle karşılaştırıldığında, etki büyüklükleri anlamlı derecede farklıydı (evlatlık-kardeş ve evlatlık-ebeveyn modelleri için p değerleri sırasıyla 0.0035 ve 0.0119). Eğer tahminleri etrafında bir simetri varsayarsak, C varyansı tahminleri çok hassas olmadığı için bu durum her iki model için de artık geçerli olmayacaktır.
Bunun bir açıklaması, ikiz ve biyolojik kardeş modellerinde C'nin derecesini yukarı doğru çıkartan ama evlatlık modellerinde bunu yapmayan eş seçimi (assortative mating) olabilir. Eş seçimi katsayısı 0.40 ile oldukça büyüktür ama "rastgele eşleşme" varsayımı bir kenara bırakılsa bile, ölçümler üzerindeki etki küçüktü ve evlatlık tasarımları ile ikiz/kardeş modelleri arasındaki fark, istatistiksel olarak anlamsız hale gelse de varlığını korudu.
Bu beni yazının başında bahsettiğim başka bir soruna geri getiriyor: İstatistiksel güç genellikle çok düşüktür. Bu yapısal eşitlik modellerinden elde edilen güven aralıkları çoğu zaman çok geniş olduğu için, tasarımların C'nin düzeyi konusunda uyuşmazlık yaşayıp yaşamadığını tespit etmek zordur. Frisell vd. (2011) nin 1.521.066 çiftten oluşan devasa biyolojik kardeş modelinde bile C üzerindeki güven aralığı %9 ile %16 arasında değişiyordu; 36.877 çiftlik ikiz modelinde ise ölçüm anlamsızdı. Küçük etkiler büyük önem taşıdığı için, tasarımların burada resmedilen küçük miktarlarda uyuşmazlık yaşayıp yaşamadığını bilmek potansiyel olarak önemlidir.
Kendler vd.’nin (2019) bir başka makalesinde, C tahminlerinin ikizler için üvey ve yarı kardeşlere kıyasla daha yüksek olduğu bulunmuştur. Bu oldukça mantıklı görünüyor: İkizlere özgü etkiler; dar jenerasyon etkilerini, residual/kalıntı yaş etkilerini vb. içerir. İkizler, sadece aynı yaşta olmaları nedeniyle bile, ikiz olmayanlara göre çevresel nedenlerle birbirlerine daha benzer olmalıdır. Çünkü büyük örneklemler olmadan yapılan yaş düzeltmeleri, gelişim süreci devam eden bireylerden oluşan örneklemlerde (çoğu çocuk örnekleminde olduğu gibi) kolaylıkla kalıntı yaş etkileri bırakabilir.
Dolayısıyla bu örneklemde, üvey ve yarı kardeşler için C'nin daha düşük olmasının, çeşitli nedenlerle büyüme sürecindeki çevre paylaşımının gerçekten azalmasından kaynaklandığını düşünüyorum. Bunun C'nin nasıl işlediği ve düzeyinin ne olduğu konusundaki anlamı hala belirsizdir. Örneğin, ikizlere özgü etkiler ile kardeşlere özgü etkiler, etkilenen bilişsel beceriler açısından farklıysa ne olur? Bunların hiçbirine henüz net cevaplarımız yok.
Yaşın rolü de hala tamamen netleşmiş değil. Yaş ilerledikçe, biyolojik kardeşlerin ve çift yumurta ikizlerinin birbirine çok benzer hale geldiğini biliyoruz. IB'nin alıntıladığı Cesarini ve Visscher'ın çok geniş ölçekli verilerinde, yetişkinlikteki özellik ölçümleri için farklar küçüktür ama "küçük" ifadesi yanıltıcı olabilir: Akrabalar arası korelasyonlar, çift yumurta ikizleri için (doğum öncesi etkilerden olumsuz etkilenmiş olabilecek boy uzunluğu hariç), birlikte büyüyen biyolojik kardeşlere göre tutarlı bir şekilde biraz daha yüksektir.
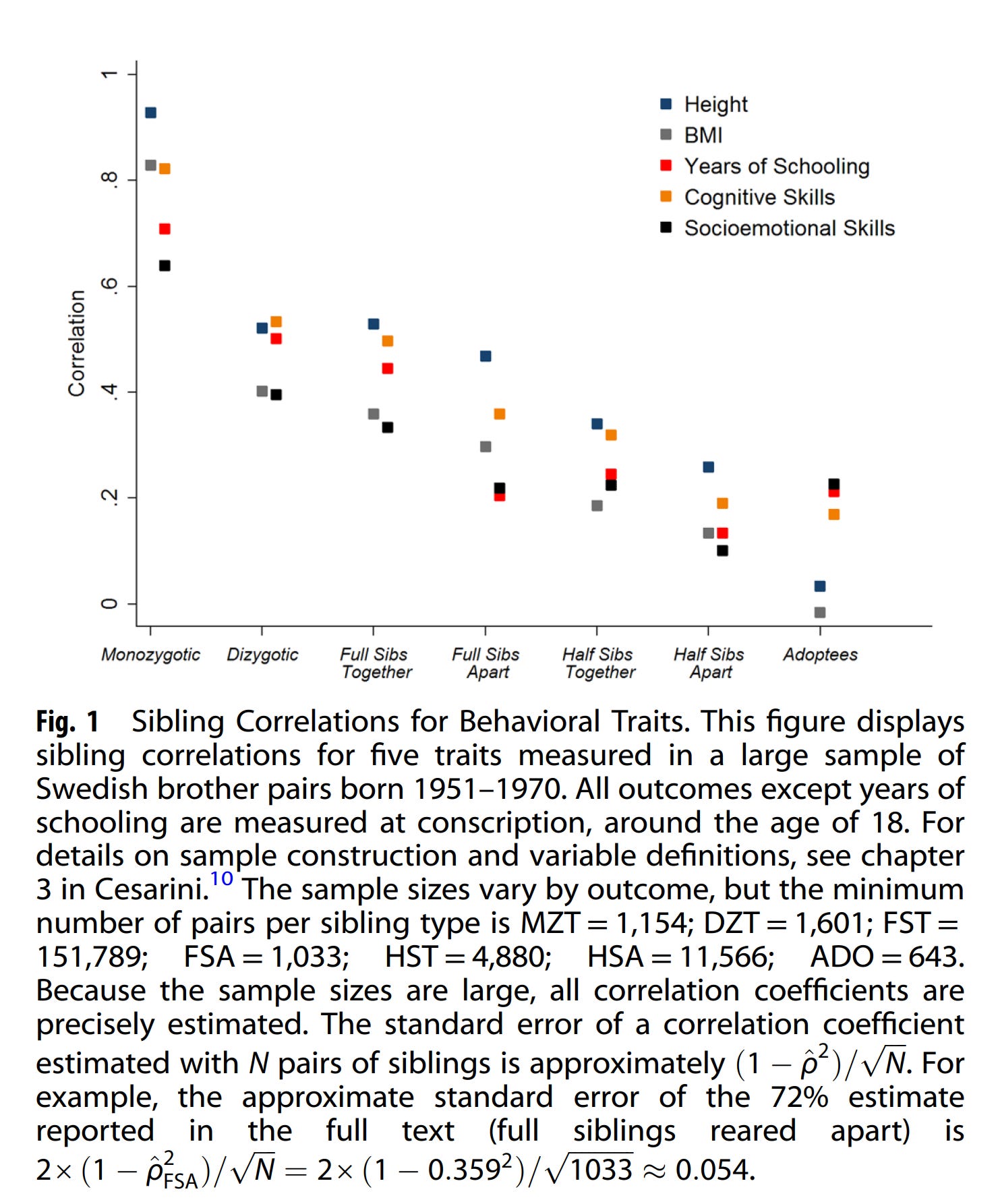
Çift yumurta ikizlerinin benzerliğinin yaşla birlikte azaldığını, Scarr-Rowe etkisinin C'nin sönümlenmesi (fadeout) yoluyla açıklandığını biliyoruz. Hatta bu durumun, 1980'lerin sonu ve 1990'larda Wilson Etkisi tam olarak anlaşılmadan önce, A'nın (genetik etki) diğerlerinin bildirdiğinden daha düşük olabileceğine dair bazı tartışmaları açıkladığını da biliyoruz. İlerleyen çalışmalarda, yaşlanmanın, örneğin ikizler yerine biyolojik kardeşlerin olduğu örneklemlerdeki etkileşim etkilerini, anlamak açısından başka şekillerde önem taşıyıp taşımadığını belirlemek, mümkün olabilir.
Büyük yaş farkı olan kardeşlerin fenotipleri başka nedenlerle de daha az kıyaslanabilir olabilir. Örneğin, kardeşler aynı yaşlarda ama yıllar arayla test edilirlerse, farklı IQ testlerine tabi tutulabilirler veya Flynn etkisi puanları etkileyerek sonraki kardeşin puanını yapay nedenlerle artırabilir veya azaltabilir. Ayrıca kardeşlerin ikizlere göre tamamen farklı okullara gitme olasılığı daha yüksektir, bu da kardeş karşılaştırmalarında daha fazla varyansa yol açabilir. İkizlerde bu tür sorunlar yoktur ve kardeş verilerinde bu sorunları düzeltmenin HER zaman işe yarayan kolay bir yolu yoktur.
Sonuç
Paylaşılan çevrenin (C) doğası hakkında anladığımız pek çok şey olduğu gibi, hala anlamadığımız pek şey de mevcut. IB, popüler ama bilimsel dayanağı zayıf olan "C=0" (paylaşılan çevrenin etkisi yoktur) önermesine karşı kanıtları öne çıkararak gayet iyi bir şey fakat akademik olarak C ile ilgili öğrenilecek çok şey var. C'nin ne kadar büyük olduğunu, duruma göre nasıl değiştiğini, yaş ve ölçüm kalitesinden nasıl etkilendiğini, tam olarak hangi çevresel unsurlara atıfta bulunduğunu, hangi fenotipler ve senaryolar için geçerli olduğunu ve daha fazlasını ortaya koyacak yeni çalışmalara ihtiyacımız var. Eğer C üzerinde bir müdahalede bulunmak ya da C varyansını artırmak veya azaltmak istiyorsak, bu tür bilgilere ihtiyacımız olacak.